

The biggest mistake beginners often make it over-supplementing their tortoises with excess calcium powders, when all they really need is a cuttlebone.
For calcium supplementation, we always offer a cuttlebone in each enclosure – that’s it! Whether they want to use it or not is their choice. The reasons for which we do not over supplement our tortoise with calcium powders is because tortoises excrete excess calcium via urine and overcharging their urinary excretion system with calcium can cause uroliths or bladder stones (Kamphues, 1991). Many beginners, and often veterinarians, justify over supplementing tortoises with calcium because they will, “just excrete the excess in their urine.” Although this is true, there is more to consider…
Liesegang et al. (2001) has shown that tortoises do excrete excess calcium in their urine. However, studies also show that the tendency to excrete calcium via urine is frequently the cause of uroliths or bladder stones (Kamphues, 1991). Why have these animals evolved this way if it causes negative effects, such as bladder stones?

Sulcatas are very prone to baldder stones. Although their larger anatomical size means stones are less likely to develop into a clinical emergency, it can still result in death. If excess calcium is presented in large quantities over time, bladder stones grow to sizes that can no longer be passed. These stones need to be surgically removed (see below). Here, our rescue sulcata is pictured with a few of the several stones he has passed upon surrender. With a change in diet, the stones should not continue to grow and we will hope he will be able to pass all stones that he still retains.
This tortoise’s case shows just how large bladder stones can become. This tortoise was not rescued by us, but by Ashley Ostrander and her family in New York. After owning the tortoise for only 4 weeks, they chose to undergo expensive, invasive, but very necessary surgery to remove the stone (pictured next to tortoise)! This stone developed over many years due to poor care from the previous owners. This tortoise is fortunate it survived.
Tortoises are hindgut-fermenters. This means that micro-organisms thrive downstream their small intestines, and aid in digestion of organic material and cellulose. In a natural environment, herbivorious tortoises will graze exclusively on plant material. Calcium in plant material is tightly bound to the plant cell-wall and is unavailable for absorption until reaching the hindgut, where microbes ligate (or release) the calcium. Therefore, in the natural environment, these tortoises rarely experience high levels of calcium in their small intestines – which would otherwise be excreted in their urinary tract. With a natural diet, they are rarely in danger of overcharging their urinary excretion system with excess calcium (Leon et al., 1979). [This is in contradictory to foregut-fermenting animals which have developed feedback mechanisms to avoid high calcium levels in their urine and prevent precipitates as uroliths.] If we provide a tortoise with an unnatural amount of readily available calcium in their small intestines, the result will undoubtedly be bladder stones and potential death. Hindgut-fermenters never needed to evolve such mechanisms because they should never experience high calcium availability in the small intestine – so think twice before you dump highly available calcium (in liquid or powder) on their food! These animals’ digestive systems are not designed to handle this.
But calcium is still very important part of a tortoise’s diet, so if dumping calicum on their food is not a good idea, how do we ensure they are staying healthy? The key is in the Calcium to Phosphorous ratios (Ca:P). As we said above, microbes are very important to the digestion process in hindgut-fermenters. In fact, the microbes are responsible for the digestion and breakdown of organic matter and cellulose, as well as the production of volatile fatty acids – the main energy source of herbivores (Breves et al., 1987; Komisarczuk et al., 1987). Micro-organisms, by dry weight, are 0.1-0.3 Ca/kg (Durand et al., 1988) and 6-19 P/kg (Komisarczuk et al., 1987). Therefore, Phosphorous becomes very important for maintaining healthy bacterial growth in the hindgut. Hindgut-fermenters have likely developed a mechanism to remove or absorb calcium from their food to maintain low Ca:P ratios for ideal bacterial conditions. Thus, if their food has a high Ca:P ratio, they will experience higher percentages of calcium absorption and use. This is confirmed through studies in horses that show a natural diet of roughage leads to a higher calcium use as compared to horses on concentrated calcium pre-prepared mixes or foods consisting of pre-ground plant material (Stadermann et al., 1992). The same conclusion has been found with other hindgut-fermenters as well (Leon et al., 1979; Clauss et al., 2003).
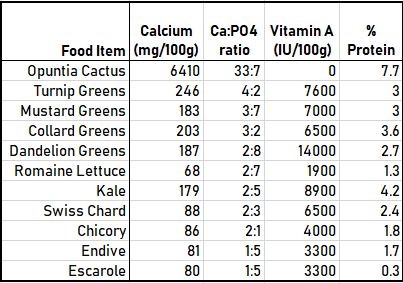
It may seem difficult to think about monitoring their diets for appropriate Ca:P ratios, but it doesn’t have to be a precise value. It is often stated that an ideal ratio of dietary calcium to phosphorous for reptiles is somewhere between 1:1 and 2:1 (Frye, 1991). The natural diet of the greek tortoise, Testudo graeca, has a ratio between 4:1 and 5:1 (Highfield, 1990). In this case, we should aim for anything with a Ca:P ratio 1:1 and above. Simply by learning which greens have better ratios than others is a great place to start (See Figure 1). For example, our tortoises love to eat Endive and Escarole, but we always make sure to add Turnip and collard greens to the mix too. Finding a reliable source of Opuntia Cactus is very beneficial to boost the ratio as well. In the Midwest, it can often be found at Mexican Food Markets at very reasonable prices.
For more advanced learning on this topic, the most informative article I have found can be accessed here: https://www.researchgate.net/profile/Marcus_Clauss2/publication/50913876_Getting_it_out_of_the_digestive_system_hindgut_fermenters_and_calcium/links/552d7cc10cf2e089a3ad776a.pdf
References:
Breves, G. et al. (1987) Effects of dietary phosphorus depletion in sheep on dry matter and organic matter digestibility. J. Anim. Physiol. Anim. Nutr. 58: 281-286.
Clauss, M, et al. (2003) Studies on feed digestibilities in captive Asican elephants (Elephas maximus). J. Anim. Physiol. Anim. Nutr. 87: 160-173.
Durand, M. et al. (1988) Influence of major minerals on rumen microbiota. J. Nutr. 118: 249-260.
Frye, F. 1991. Biomedical and Surgical A spects o f C aptive R eptile H usbandry. Krieger Publishing Co. Malabar, Florida.
Highfield, A. 1990. Keeping and Breeding Tortoises in Captivity. England, R and A Publishing Limited.
Innis, C. (1994) Considerations for Formulating Captive Tortoise Diets. Bulletin of the Association of Reptilian and Amphibian, Vol. 4, No. 1.
Kamphues, J. et al. (1991) Calcium metabolism of rabbits as an etiological factor for urolithiasis. J. Nutr. 121: S95-S96.
Leon, B. et al. (1979) Calcium, phosphorus and magnesium excretion in the rock hyrax. Comp. Biochem. Physiol. A. 64: 67-72.
Liesegang, A. et al. (2001) Influence of different dietary calcium levels on the digestibility of Ca, Mg, and in captive-born juvenile Galapagos giant tortoises (Geochelone nigra). Zoo Biol. 20: 367-374.
Komisarczuk, S., et al. (1987) Effects of phosphorus deficiency on rumen microbial activity associated with the solid and liquid phase of a fermentor (RUSITECH). Repro. Nutr. Dev. 27: 907-919.
Stadermann, B. et al. (1992) Calcium and Magnesium absorption. bei Rauhfutter oder Mischfuffer. Pferdeheilkunde. Special Issue 1: 77-80.
Top of Form