For herbivorious tortoises, excess protein is metabolized and excreted as insoluble uric acid (urates). Urates are not a result from excess calcium in the diet, nor are they a result from being dehydrated, which are both commonly cited by beginners as the cause of urates. In reality, the presence of urates indicates elevated levels of protein in the diet. The consistency of the urates indicates the level of hydration. Calcium intake is completely unrelated to urate production.
For herbivorious tortoises, limiting protein intake is very important. If uric acid (urates) levels rise beyond what can be flushed by the kidneys, uric acid crystals can be deposited in joints and vital organs. This can lead to articular or visceral gout (Frye, 1991). Additionally, excess protein can also be converted by the liver to carbohydrate and fat, leading to liver disease (Highfield, 1990; Jarchow, 1992).
The tables are turned when we are discussing omnivorious tortoises, however. Any forest-dwelling species, such as redfoots, yellowfoots, elongated, or hingebacks, are from native regions that are plentiful of worms, millipedes, slugs, fungi, and other protein rich food sources. Tortoises, being opportunistic in nature, will accept any food sources. Therefore, these tortoises have evolved different digestive systems that can tolerate higher protein diets. In fact, feeding these tortoise species a protein source every 1-2 weeks is a healthy option, especially because animal proteins are an additional source of vitamin D (making them less reliant on UV exposure).
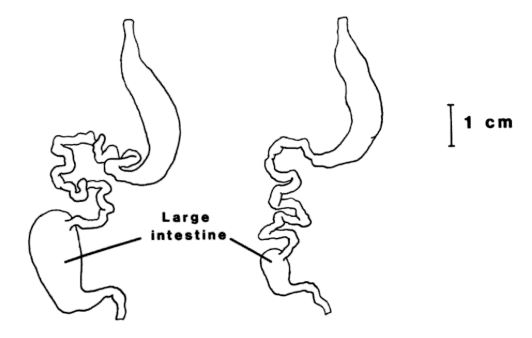
Herbivorious tortoises are not at a complete disadvantage as compared to their omnivorious relatives, however. There is a general trend for herbivorous species to have longer digestive tracts than do omnivorous species (Troyer, 1991). Digestive tracts, illustrated below from Troyer (1991), display the key differences in digestive systems of herbivorious reptiles (left) and omnivorious reptiles (right). The key difference is that herbivorious reptiles are hindgut-fermenters and this microbial fermentation system that degrades plant fiber is housed in the “cecum,” which is the upper portion of the large intestine (Guard, 1980). This results in a larger and longer large intestine in herbivorious tortoises. This alteration in the digestive system actually gives herbivorious tortoises an advantage in a completely plant-based diet. Herbivorious tortoises are more efficient (nutrient or energy gain per unit) than omnivorious tortoises (Bjorndal and Bolten, 1993). Understanding these differences is important to consider when developing a feeding regimen for your tortoise.
References:
Bjorndal, K. A., and A. B. Bolten. 1992. Body size and digestive efficiency in a herbivorous freshwater turtle: advantages of small bite size. Physiological Zoology 65:1028–1039.
Frye, F. 1991. Biomedical and Surgical A spects o f C aptive R eptile H usbandry. Krieger Publishing Co. Malabar, Florida.
Guard, C.L. (1980) In Comparative Physiology: Primitive Mammals (K. Schmidt-Nielson, L. Bolis, and C.R. Taylor, eds.), Cambridge University Press, New York, p. 43.
Highfield, A. 1990. Keeping and Breeding Tortoises in Captivity. England, R and A Publishing Limited.
Jarchow, J. 1992. Practical aspects of reptile nutrition. Regional Proceedings of the AAZPA, 272-275.
Troyer K (1991) Role of microbial cellulose degradation in reptile nutrition. In: Haigler CH, Weimer PJ, eds. Biosynthesis and Biodegradation of Cellulose, pp. 311–325. New York: Marcel Dekker.